

「繁殖生理学」という授業が4年生の後期にあり、動物の性周期や性ホルモンの動態について学びました。
性ホルモンは相互に影響を与えており、その全体像を理解するのには非常に骨が折れます。
学生だけでなく、農場で働いている方々にとってもこれを理解することは非常に重要であると思います。
勉強しているときに「パッと見て分かりやすいイラストがあれば良いのに…」と思っていたので、自分で図を作ってみました。
今回の記事ではそれらの図を使って、できるだけ分かりやすく牛の繁殖生理について解説していきます!
目次(移動できます)
発情周期とは?
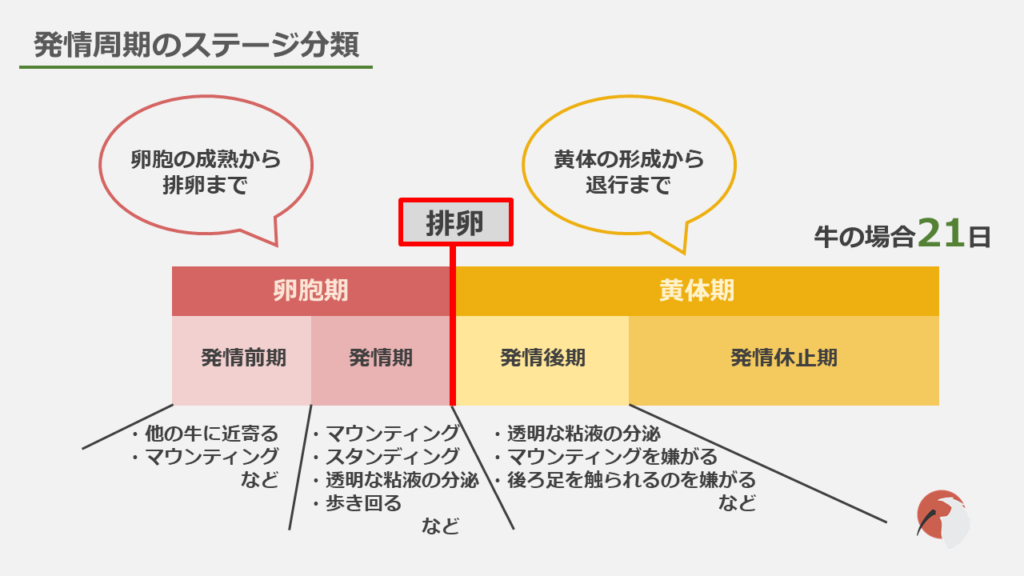
ホルモンの動態について書いていく前に、発情周期についてご説明したいと思います。
発情周期とは言葉の通り、繁殖活動期のメスで発情が起こる周期のことです。
これは排卵日から次の排卵日までの長さと同じです。
発情周期は黄体期の有無により、完全発情周期と不完全発情周期に分けられますが、牛をはじめ多くの哺乳類は排卵後に黄体期が見られる完全発情周期を示します。
発情周期は、「卵胞・黄体」と「発情」の2つの視点から見ることができます。
まずは「卵胞・黄体」の視点からです。
完全発情周期では、卵胞発育、排卵、黄体形成、黄体退行が周期的に繰り返されます。
卵胞の成熟から排卵までの期間を卵胞期、黄体の形成から退行するまでの期間を黄体期と呼びます。
次に「発情」の視点から見ると、完全発情周期は、発情前期、発情期、発情後期、発情休止期の4つのステージに分類されます。
発情期は交尾に最も適した時期で、乗駕を許容するスタンディングやマウンティング、他にも外陰部から粘液を出したり歩数が増えるなどの変化が見られます。
排卵後に黄体期が見られないものを不完全発情周期と言い、マウス、ラット、ハムスターなどがこれに該当します。
これらの動物は黄体の生存に不可欠であるプロジェステロンを不活化する酵素を持つために黄体が存続できません。
しかし、交尾が行われるとこの酵素の産生が抑制されるため、黄体が形成されます。
牛の発情周期

牛は1年を通して繁殖活動を行うことができる周年繁殖で、妊娠しない限り1繁殖期に発情周期を繰り返すことができる多発情動物です。
発情周期の長さは経産牛では平均21日、未経産牛では少し短く、平均して20日です。
周年繁殖動物は牛の他に豚や犬が該当します。
余談ですが、周年繁殖動物とは逆に、生殖活動が可能な季節が決まっている動物を季節繁殖動物と言い、猫、馬、羊、山羊がこれに当たります。
また、1繁殖期に発情周期が1回しか見られない動物を単発情動物といい、犬がその好例です。
卵胞の発育と退行
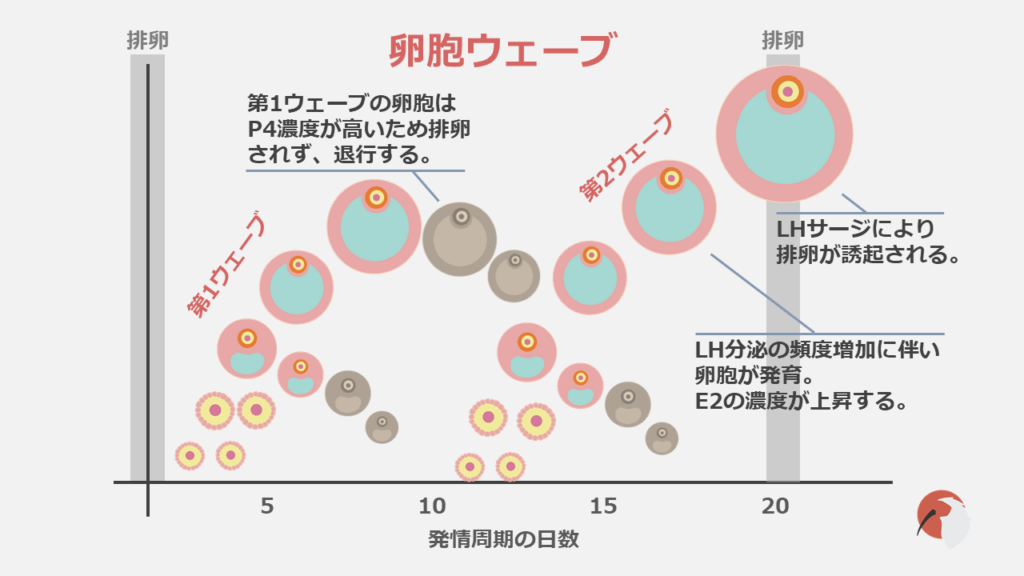
牛では1回の発情周期中に2~3回の卵胞群の発育が観察されます。
この卵胞群の発育は卵胞ウェーブと呼ばれます。
卵胞ウェーブは多数の小卵胞の出現から始まります。これは卵胞の動員と呼ばれます。
その次に多数の小卵胞の中から選ばれた1個の卵胞(主席卵胞)は発育し、成熟します。選ばれなかった卵胞たちは退行します。
これは卵胞の選抜と呼ばれます。卵胞の選抜の仕組みは卵胞刺激ホルモン(FSH)の章で解説してあるのでそちらをご覧ください。
上の図にもあるように排卵後初めて出現する卵胞ウェーブ(第1ウェーブ)の主席卵胞は成熟せずに閉鎖してしまいます。
これは黄体から分泌されるプロジェステロン(P4)の作用により、黄体形成ホルモン(LH)のパルス状分泌が抑制されるためです。
すると次の卵胞ウェーブ(第2ウェーブ)が出現します。
発情周期の終盤になると黄体が退行しP4が少なくなるため、発情周期中の最後の卵胞ウェーブの主席卵胞は成熟することができ、排卵に至ります。
個々のホルモンについて
それでは以下より、発情周期中のホルモンの変化について1つずつ見ていきたいと思います。
黄体形成ホルモン LH
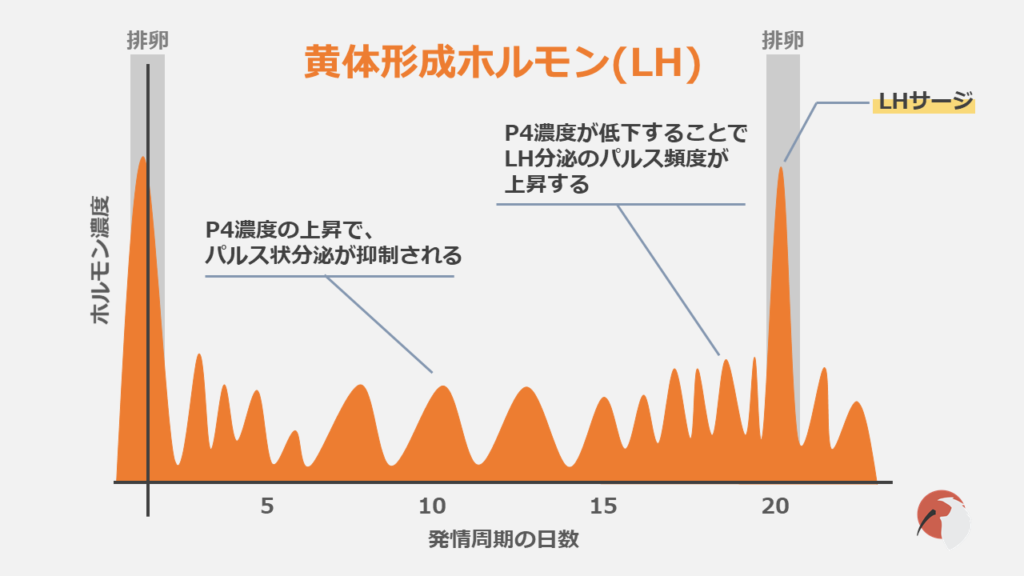
LHはFSHとともに、性腺刺激ホルモンと呼ばれ、脳下垂体前葉の性腺刺激ホルモン産生細胞(ゴナドトロフ)で合成されます。
LHの分泌頻度が上昇すると発育を開始した胞状卵胞に作用し、卵胞をさらに発育させ、それに加えて黄体の機能を維持させることもできます。
また、FSHとともにエストロジェンの合成と分泌を促進します。
LHを説明するうえで重要となってくるのが「LHサージ」です。
LHサージとは卵胞が十分に発育した時に発生する、急激なLH濃度の上昇のことです。
LHサージによって以下のことが引き起こされます。
- 排卵の誘起
- P4合成の増加
- 黄体化の開始
- 減数分裂の再開
①はその通りで、LHサージにより卵巣から卵子が放出されます。
②ではP4合成が増加することで、コラーゲンを分解するコラゲナーゼという酵素が増加します。
これにより、卵胞のコラーゲンが分解されて卵胞壁が薄くなるので排卵しやすい状態となります。
また、P4が増加することで子宮において着床性増殖が見られ、受精卵が着床しやすくなります
P4の子宮に対する作用については後述します。
③により、排卵後の卵胞が黄体になります。黄体はP4を産生するため、さらにその量が増加します。
最後に④の減数分裂の再開についてです。
排卵に至るまで、卵子は第一減数分裂前期で止まっています。その後LHサージが起こることで第一減数分裂が再開し、第一極体を放出して、第二減数分裂に進みます。
以上のことから分かるように、LHサージは単に排卵を引き起こすだけでなく、卵子の成熟にも関わる非常に重要な因子であると言えます。
卵子の形成は、メスの個体が母親のおなかの中にいる胎生期から始まります。
胎生期のうちにほとんどの哺乳類で成熟分裂に至り、そして上述したように、排卵するまでは第一減数分裂前期で停止します。
LHサージによって第二減数分裂へと進みますが、第二減数分裂中期で再度停止し、受精を待ちます。
精子が侵入すると二次極体を放出し第二減数分裂を完了して、受精卵となります。
このように卵子の成熟は、”胎生期から受精まで”という非常に長い期間にわたって行われます。
卵胞刺激ホルモン FSH
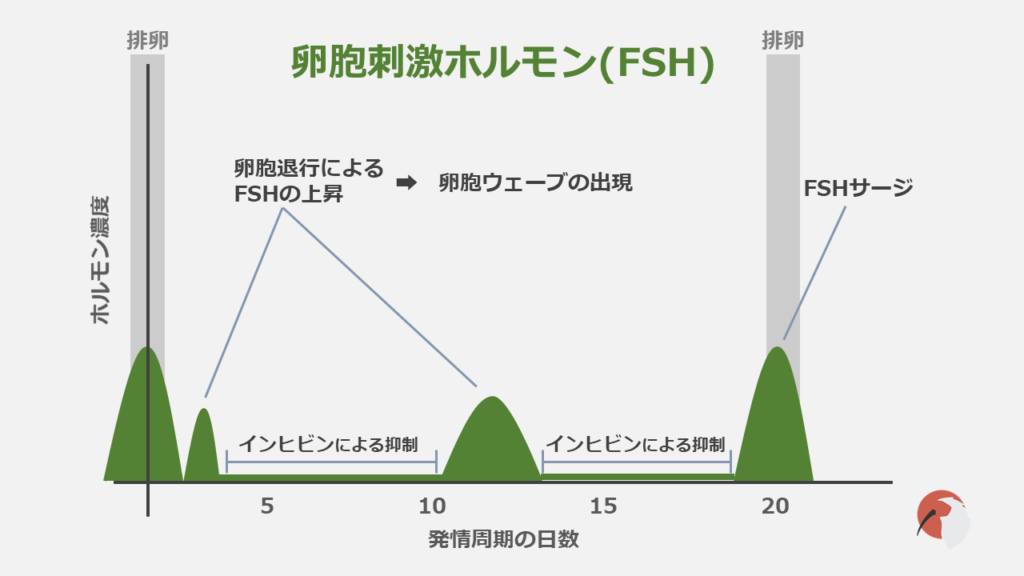
LHと同様にFSHも性腺刺激ホルモンの1種です。
メスにおいて卵胞の発育に関与しますがFSH単独では卵胞を成熟させることができず、LHとの協力が必要です。
FSHの分泌はE2とインヒビンによって抑制されますが、排卵によりこれら2つの濃度が下がるとFSH濃度の急激な上昇が観察され、新たな卵胞が出現します。
卵胞の出現と選抜もFSHによって調節されます。
FSHが上昇すると、その約2日後に新たな卵胞ウェーブが出現します。
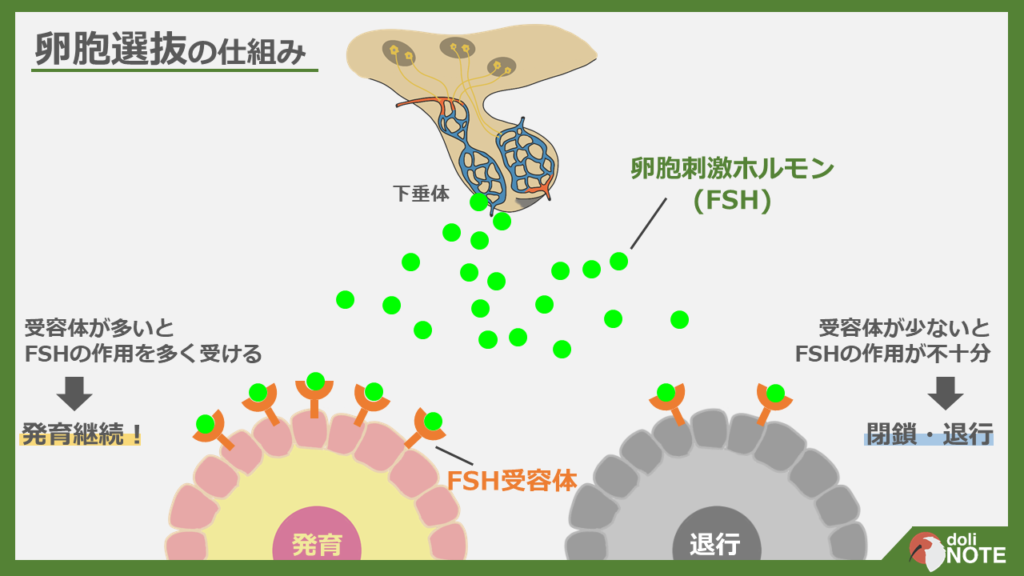
下の画像は卵胞選抜の仕組みを図示したものです(PDFはこちら)。
卵胞の表面にはFSH受容体がありますが、この数が卵胞によって異なります。
卵胞の内、FSH受容体を多く持っているものはそうでないものと比べてFSHの効果を大きく受けて発育します。
他の卵胞は発育することができずに閉鎖・退行していきます。
これがFSHが卵胞を選抜する仕組みです。
LHとFSHはともにGnRHによる制御を受けているので、LHサージが起こるとFSHでも同様な上昇が見られます。
これは一応「FSHサージ」という名前が付けられているのですが、その役割は不明な点が多く、LHサージの”おまけ”程度にしか考えられていませんでした。これだけ大きな濃度の上昇なのに教科書でも説明されていません。
しかし、FSHサージによって受精率が高まったり、卵巣が過剰に大きくなってしまう病気の発生率が低下したとの報告があるので、やはり何かしらの役割を持っていると考えられます。
エストラジオール E2
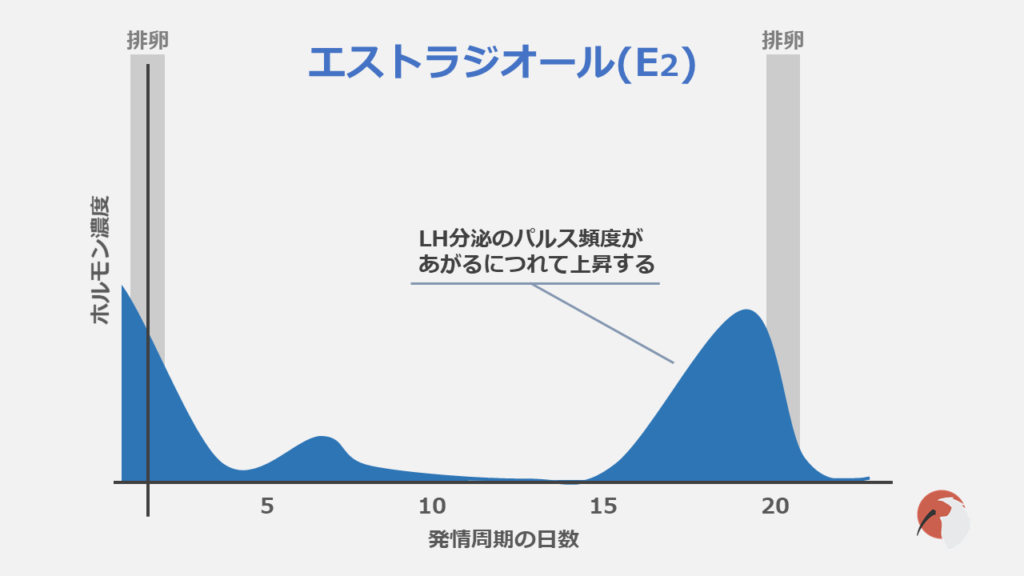
血中に存在するエストラジオールは主として卵胞の顆粒層細胞で産生されます。
発情周期中はLH分泌の頻度増加に伴いE2も上昇します。
E2が上昇するとGnRHサージが引き起こされ、GnRHサージによりLHサージが引き起こされ、排卵が誘起されます。
排卵が終わるとE2の濃度は減少しFSH分泌に対する抑制が解除され、FSHの濃度が上昇します(FSH抑制作用はインヒビンの方が強いです)。
エストロジェンは他にも性行動をつかさどる中枢機構に作用し発情を引き起こします。
また、卵管運動を亢進させるとともに子宮内膜上皮の発育増殖と粘液分泌活動などを促進します。
これは分娩の過程において非常に重要な働きです。
エストロジェン(エストロゲン)はエストロン、エストラジオール、エストリオールの3種類のホルモンの総称です。
これら3種類のホルモンは水酸化物イオン(OH-)の数が異なります。
資料によって呼び方が異なるので混乱する方がいるかもしれませんが(僕は「??」となってました)、基本的には同じものと考えて差し支えありません。
ちなみに、エストラジオールの作用が最も強いです。
プロジェステロン P4
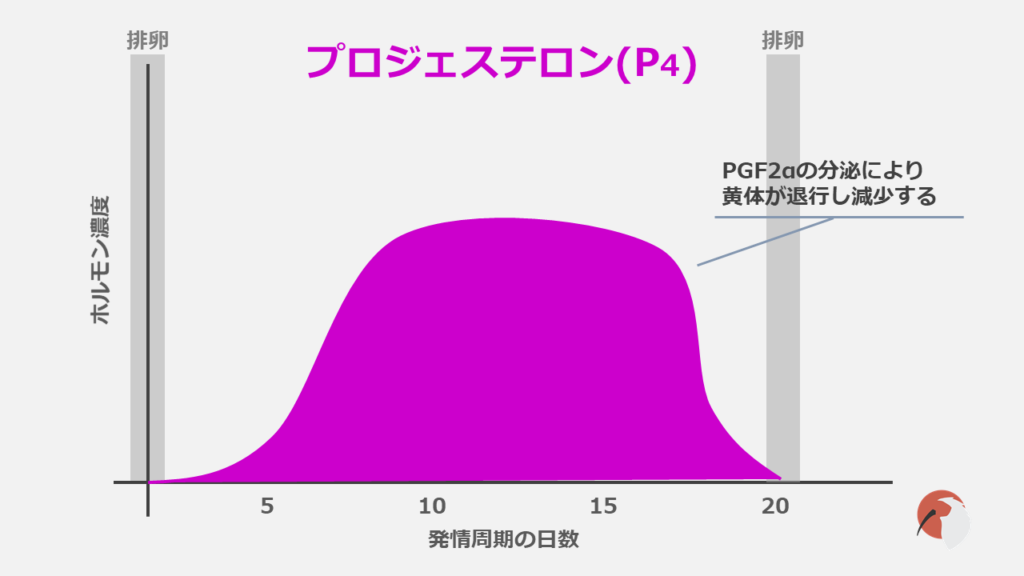
血中に存在するプロジェステロンは主として卵巣の黄体で産生されます。
妊娠を成立、維持させるために副生殖器や内分泌系に変化をもたらすことが主な働きです。
P4はGnRHの分泌を抑制し、これによりLHのパルス状分泌が抑制されます。
P4濃度はPGF2αの分泌で低下します。これはPGF2αにより黄体が退行するためです。
排卵後に黄体形成が再度始まることで、P4濃度は上昇します。
P4は子宮にも作用します。
子宮内膜上皮に作用し、内膜上皮を増殖させ浮腫状にします。これは着床性増殖と呼ばれ受精後の胚の着床にとって非常に重要です。
他にも、子宮の自発運動の抑制、オキシトシンに対する感受性の低下、子宮の平滑筋収縮の抑制、子宮内膜の分泌機能の亢進、子宮運動の抑制および頚管の収縮などの働きを持ちます。
これらは胚着床以降の妊娠維持に必須となります。
プロスタグランジンF2α PGF2α
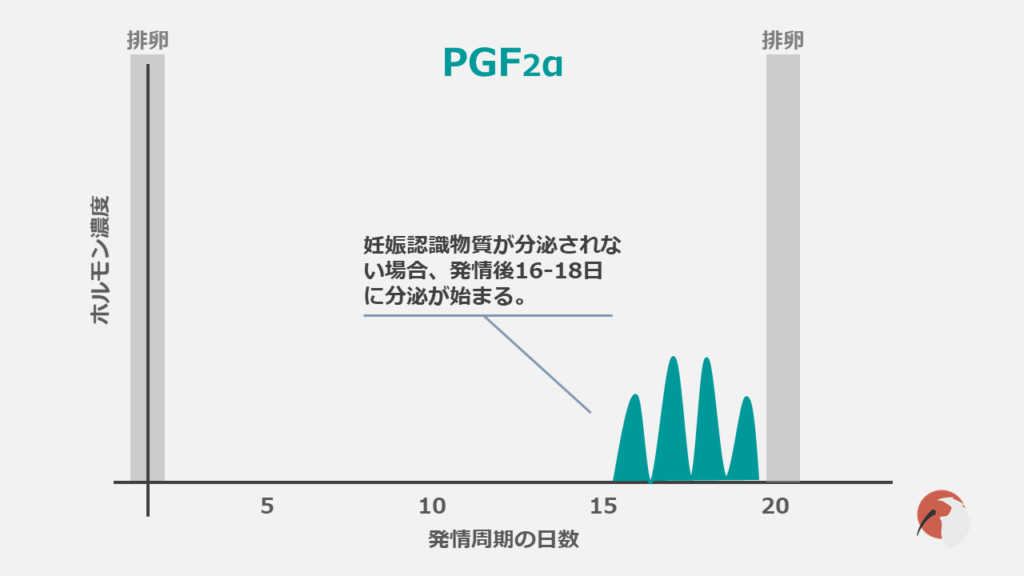
子宮内膜が主要な産生部位です。
発情後12-13日までに胚から妊娠認識物質が分泌されない場合に分泌が始まります。
黄体を退行させプロジェステロン産生を低下させる強力な黄体退行因子です。
性腺刺激ホルモン放出ホルモン GnRH
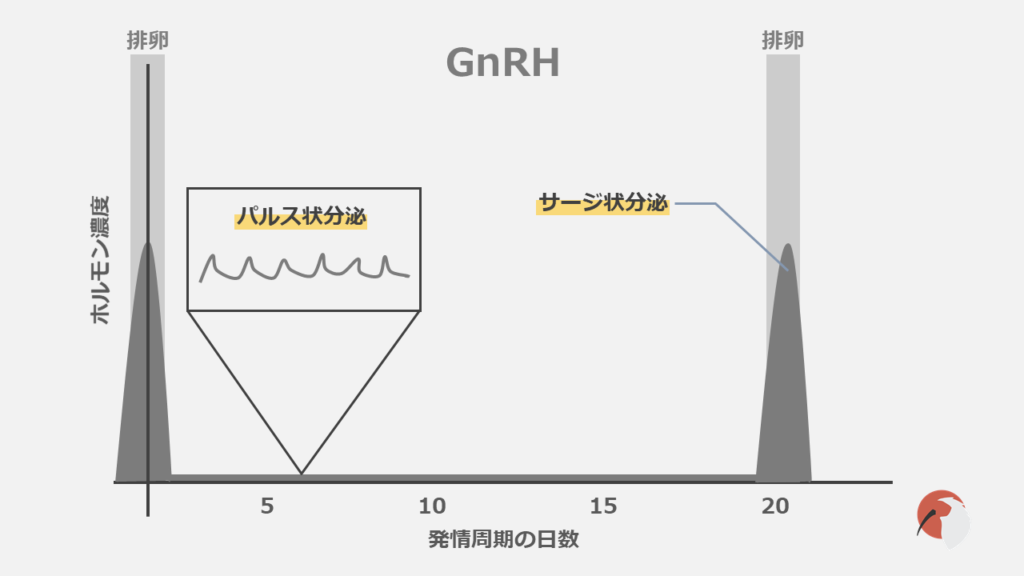
視床下部から放出されます。
下垂体前葉に作用し、LHとFSHの合成・分泌を促進します。
GnRHはパルス状分泌で、この頻度の変化が下垂体からのLHとFSH分泌に影響を及ぼしています。
このようなパルス状分泌はGnRHパルスジェネレーターによって制御されています。
また、LHサージはGnRHのサージ分泌で起こります。
これを制御するGnRHサージジェネレーターが予想されていますが明らかになっていません。
インヒビン
卵胞の顆粒層細胞から分泌されます。これはE2と同じところです。
卵胞の発育に伴って血中濃度が上昇しますが、排卵によって分泌は停止し血中濃度が減少します。
つまり、血中濃度が減少するタイミングがE2と同じです。
主な作用は下垂体前葉に作用してFSH分泌を抑制することです。
これら2つのホルモンは卵胞の顆粒層細胞で産生され、FSHの抑制作用を持ちます。
そのため、グラフでは一緒にされることが多いですが、分泌開始の時期が少し異なります。
インヒビンは卵胞発育の初期から分泌されるのに対して、E2は卵胞が成熟してから分泌されます。
一方で減少するタイミングは同じで、それは排卵時です。
すでに説明したように、この2つのホルモンの減少によりFSHの抑制が解除され、FSHサージが見られます。
1つにまとめてみました
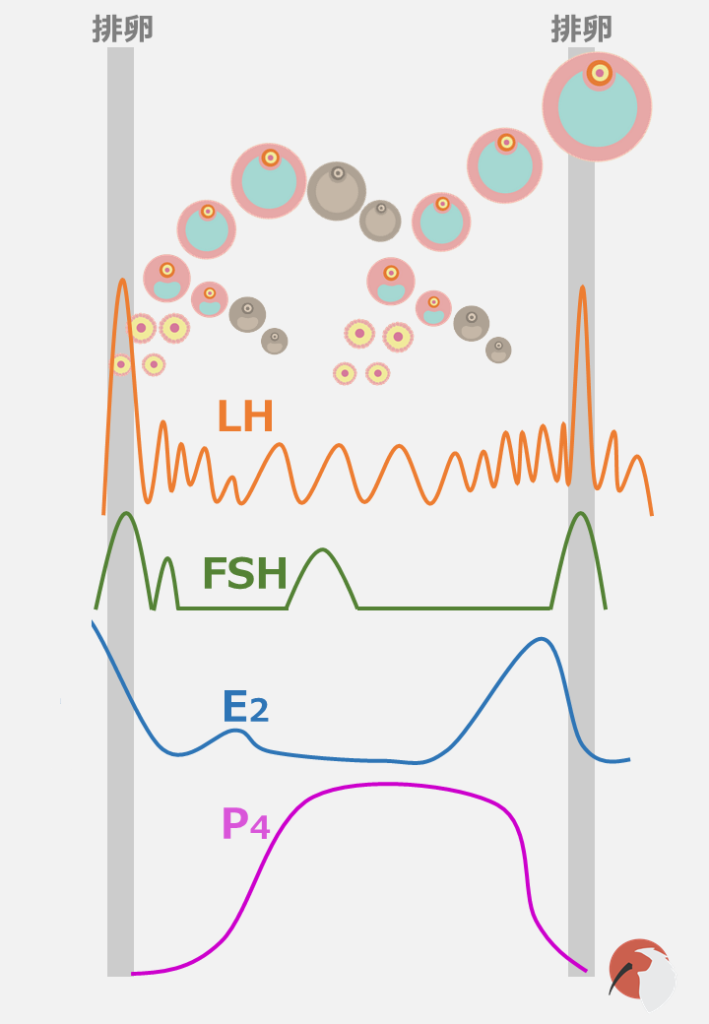
LH、FSH、E2、P4のグラフと卵胞ウェーブの図を一つにしてみました。
これを見ると、P4濃度が高い時はLHの分泌頻度が低いこと、E2の低下によってFSHが上昇していること、卵胞ウェーブがFSHの上昇で発生していること、卵胞の発育に伴いE2濃度が上がっていることなど、上で説明してきたことが一目で分かるかと思います。
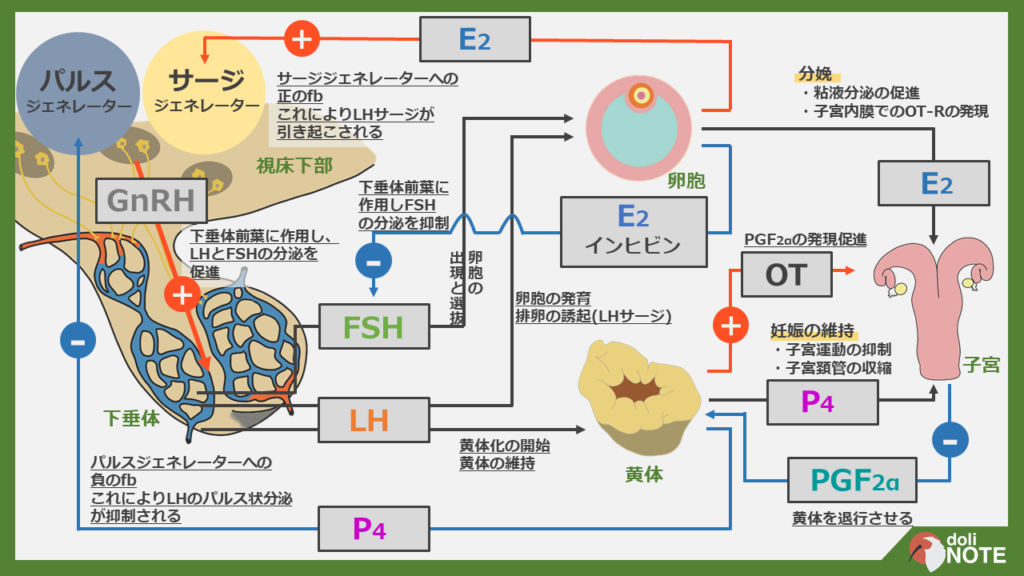
ホルモンの相互関係
上で見てきたように、ホルモンはお互いに影響を与えて、産生を調節しあっています。
この関係ががなかなか理解しづらいので図にしてみました。
こちらからPDFで細かく見ることができます。
反芻動物の黄体退行のメカニズム
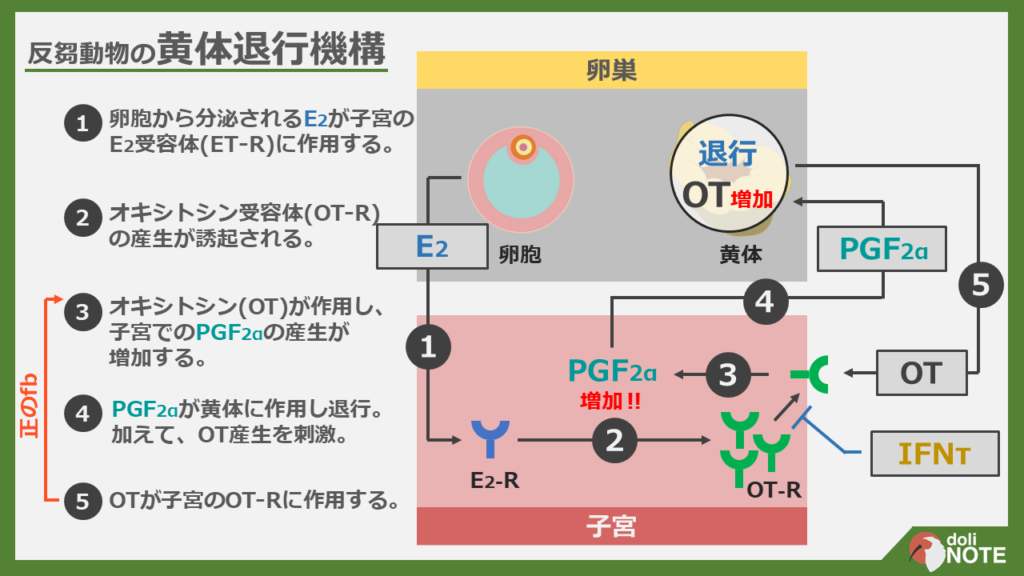
次の発情の3~5日前になっても妊娠していない場合は黄体が退行し始め、次の排卵に備えます。
黄体の退行はどのような仕組みで行われているのでしょうか?
これも図にすると分かりやすいと思ったので作成しました(PDFはこちら)。
まず排卵に至る前の卵胞に目を向けてみましょう。
そこから分泌されたE2は子宮内膜に作用して、オキシトシンレセプター(OT-R)の発現を増加させます。
排卵後に形成された黄体から分泌されたOTは子宮内膜のOT-Rに作用します。
すると、子宮内膜からのPGF2αの産生が誘起されます。
PGF2αは黄体に作用し、黄体を退行させるとともにOTの産生を刺激します。
OTがさらに分泌されたため、PGF2αの量もさらに増加し、黄体がどんどん退行していきます。
PGF2αとOTはお互いの産生を増加させあっていることが分かるかと思います。
このような関係は、「正のフィードバック」と呼ばれます。
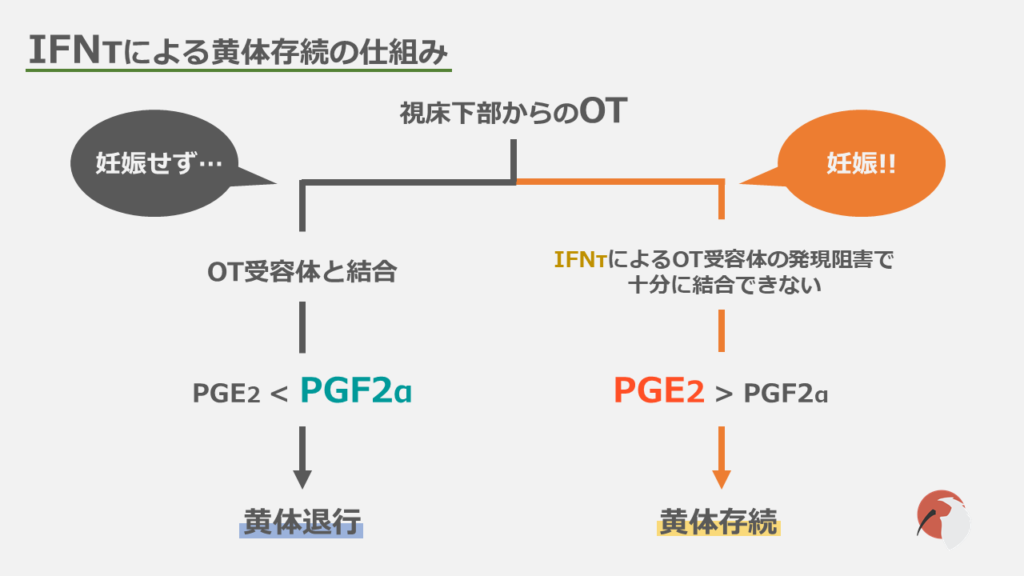
次に妊娠した場合についてです。
妊娠が成立すると胚からインターフェロンタウ(IFN-τ)が分泌されます。
IFN-τはOT-Rの発現を阻害します。
すると子宮のOTに対する感受性が低下し、それに伴いPGF2αの産生も低下するため、結果的に黄体が維持されます。
牛の発情周期から始めて、性ホルモンを一つずつ見ていき、最後に黄体退行の仕組みについて書きました。
発情周期中には様々な性ホルモンが関与するうえに、それらが複雑に影響を与え合っているので理解するのが一苦労だと思います。
僕もテスト勉強している時に同じように感じていたので、図を交えながらできるだけ分かりやすく解説したつもりです。
この記事を読んでくださった方々が「ふむふむ、そういうことかー」と理解が少しでも深まったのであれば、嬉しい限りです。
最後まで読んでいただきありがとうございました。
参考資料
・「獣医繁殖学 第4版」中尾敏彦、津曲茂久、片桐成二 文永堂出版 2019年
・「インヒビンとエストラジオールによる視床下部・下垂体・卵巣軸の調節機構」田谷一善 日本生食内分泌学会雑誌(2003)8: 11-17

